Pick from the Past
Natural History, November 1958
![]()
Darwin’s Worms
![]()
In his last years, the great man studied annelid “intelligence”:
the questions he raised have engaged a generation of biologists.
By Georg Zappler
![]()
Mr. Zappler was born in Austria and holds a Master’s degree in Zoology from
Columbia University. He is a regular contributor of articles on the biological sciences.
![]()
| Charles Darwin’s last major publication, The Formation of Vegetable Mould, through the Action of Worms, with Observations on their Habits (1881), appeared a year before his death. This report of his study—and its challenge to later investigators—was published to honor the naturalist’s centennial year. |
CHARLES DARWIN’S NAME has become so closely linked with the theory of evolution over the past century that it is often forgotten that this same man was not only a great theoretician but also a keen naturalist—interested in a hundred different aspects of the world around him. It is fitting, during a year that sees celebration of his evolutionary hypothesis,
|
||
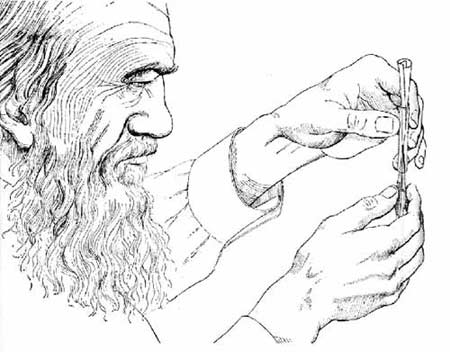
It was in the course of investigating the role that earthworms play in the formation of vegetable mold that Darwin undertook a study of this humble, but significant, invertebrate’s behavior. He noticed that, at night, these annelids (known scientifically as Lumbricus) would seize and drag leaves, petioles, pine needles and other small bits of vegetable trash into the opening of their burrows. The end result of these labors not only constituted very efficient plugging-up of the worms’ domiciles, but provided a lining for the front portion of their passageways to the surface.
If one looks for any advantages accruing to the earthworm as a result of this behavior, one is left with a number of possible choices. The most obvious benefit would be protection for the earthworm from such sharp-eyed predators as the traditional “early birds,” since the worm rests just below the hidden opening of its burrow. Another, less obvious, advantage might be a certain amount of insulation—obtained from the retention of heat produced by the blanketing, leafy plug. Possibly, a flood prevention factor may also be involved in this plugging operation. In addition to all these protective values, the nutritional factor involved in having a supply of leaves—at hand in the pantry, so to speak—should not be overlooked, for certain of the vegetation that Lumbricus drags into its burrow definitely serves as food.
To the investigator of behavior, however, the most interesting aspect of this plugging activity is the manner of execution. How does the worm go about transporting such relatively bulky objects as leaves? In what manner are these anything-but-plug-shaped objects effectively pulled into the narrow opening of the burrow? Darwin found that about eighty per cent of the withered leaves taken from the surface by earthworms had been drawn in by the tips—obviously the most convenient method when a wide object, pointed at one end, has to be drawn into a narrow tunnel. Equal in efficiency was the way in which pine needles were handled. These evergreen leaves, each consisting of two thin, pointed needles joined at the bottom, are almost always pulled in base-foremost—evidently, the most practical procedure.
At this point, any scientist—and certainly one of Darwin’s stature—starts searching for some explanation of this seemingly purposeful behavior. The searcher is faced With three conclusions: the behavior is a matter of chance, of blind “instinct” or of intelligence. Only resort to experimentation can eliminate one or more of these possibilities from consideration.
In Darwin’s case, he proceeded to set up a “controlled” situation to help him decide which was the answer. Was Lumbricus a creature that dragged leaves into its burrow willy-nilly? Or did it possess a demonstrable ability to drag in each leaf in the most practical manner? If the latter, was this an “instinctive” or inherited talent; or was it instead, a process that somehow involved reason and the capacity to learn?
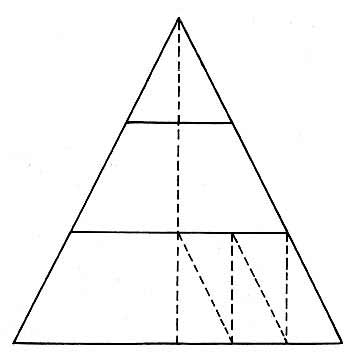
Darwin’s classic experiment, aimed at answering these questions, consisted of providing earthworms with three-inch-high paper triangles, instead of leaves. Each triangle had a pair of lines ruled on it at equal distances from, and parallel to, the base, permitting Darwin’s data to be grouped into three convenient categories: (a) triangles that had been seized and drawn in near the apex, (b) those tackled by the worms at a point along the base, and (c) the ones seized somewhere in the median area between the two ruled lines.
 Worm’s work in pulling vegetable trash into burrows, as analyzed by Darwin, showed pine needles usually seized at base; laburnum leaves about equally drawn in by base or tip; lime leaves usually taken by tip. In case of paper triangles, nearly two-thirds were seized by the tip. |
To prejudge the work involved for the worm in each case, Darwin used a pair of forceps, with which he seized the paper triangles at various points and then drew them into a tube approximately the diameter of a worm burrow (see illustration at top of page). He found that, if seized by the apex, the paper was drawn straight into the tube with its margins uncreased—forming a regularly-shaped cone. If the point of seizure was some distance below the apex, that much of the triangle would be doubled back within the tube. Such was also the case with seizure at the base, or at basal angles, although the resistance offered in these cases was, as could be expected, much greater.
Darwin reasoned that, if the worms’ actions were determined by chance, then (since the area of the basal portion of each paper triangle was five times that of the apical region) the number of triangles drawn in by the base should maintain at least a ratio of five to one over those pulled down by the apex. In addition, the basal portion offered two convenient points of attachment at its angles, compared with only one such point at the apex; also the marginal length available for seizure along the bottom part is greater than in the uppermost region. All these factors should favor the probability of finding the triangle pulled down by base rather than by apex.
| ||
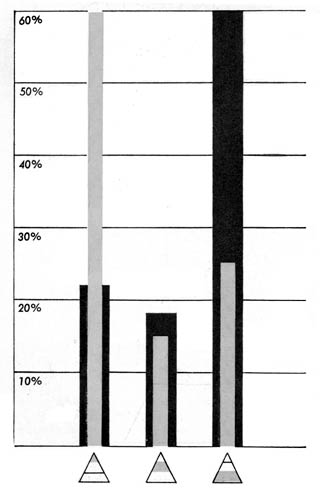
Darwin’s experimental findings, however, completely contradicted these calculations based on chance. He scattered his paper triangles on the ground near worm barrows from which leaves and other objects had been removed. Altogether, over a period of many successive nights, about three hundred triangles were drawn in by worms. Of these, more than sixty per cent had been pulled down near the apex, some fifteen per cent around the middle, and less than twenty-five per cent by the basal portion. Since nearly three times as many “leaves” had been drawn in by the apex than by either middle or base—an action that had the odds at least five to one against it—Darwin concluded that the worms’ manner of handling these objects owed little, if anything, to chance.
By now, the reader, schooled in modern experimental techniques, will be ready to point out a flaw in Darwin’s analysis. How could Darwin, having examined only the end result of the worms’ successful labors, tell whether his triangles had been initially tackled at the apex or elsewhere—with the worms "shifting their grip" in the latter cases? To put it another way, where was the observer while the worms were at work? The analysis of end results only—by modern standards—was valueless.
Darwin, of course, had not been naïve enough to overlook this serious problem. From his observation of worms kept in pots indoors, he noticed that, when the triangles were drawn in by the apex, the basal portions remained clean and uncrumpled. When a triangle had been pulled down near its bottom, however, that portion would be very much crumpled and covered with slimy dirt. Darwin reasoned, therefore, that any triangle found drawn in at the apex (and not stained or creased in the basal area) could be
|
||
He dismissed instinct for a number of reasons. In his study of worms in their natural settings, Darwin observed some striking differences in the way various kinds of leaves were handled by the annelids. The broad-based leaves of lime trees (not an endemic English species) were almost exclusively found drawn in by the apex. With laburnum leaves (almost equally pointed at both ends), however, only a little more than half were pulled down tip-first. Pine needles, as already mentioned, were almost always drawn down by the joined base: indeed, the worms continued to use the base-first method even when the tips of the twin needles were cemented or tied together by fine thread. Such a variety of reactions to different types of material led Darwin to the conclusion that instinct was not a determining factor: it seemed improbable to him that earthworms should have such a range of specialized instincts for different situations. Further, he asked himself how “English” annelids could have developed hereditary innate behavior in reference to objects totally unknown to their ancestors and so various as man-made triangles and the leaves of introduced foreign plants. Darwin’s alternative explanation was to credit Lumbricus with some degree of intelligence.
Darwin’scase for some sort of reasoning and learning in the earthworm remained fairly well established until the turn of the century, a time when students of behavior—carried away by the discovery of reflex responses—were prepared to ascribe to these newly discovered mechanisms all the functions previously considered “psychic” in the behavior of animals. As with so many creatures, such was the case with the worms.
In 1904. Elise Hanel, the German zoologist, ran a study on the leaf-pulling behavior of Lumbricus along lines similar to Darwin’s, and obtained very similar results. However, her interpretation of the findings was quite different. Like Darwin, she dismissed instinct. Such behavior, she reasoned (not very logically), could not be inherited since it had first to he acquired—acquired traits, by definition, not being transmitted hereditarily. She also denied the possibility of associative learning or learning through experience in as lowly an animal form as this invertebrate plowman. Having dismissed both these possibilities, she interpreted her data along the lines of the period’s prejudice—namely, a purely automatic response on the earthworm’s part to specific stimuli provided by the objects encountered in its wanderings.
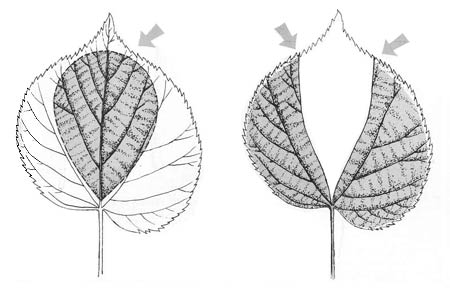
Hanel’s theoretic approach may not have been purely objective, but her experimental procedure was ingenious. She cut lime tree leaves into different shapes. In some, the distal end was made broad and rounded and the basal end narrow. In others, the leaf was split and a portion
| ||
| |

Hanel did not stop with altered lime leaves. She went on to test paper shapes—as Darwin had done—but shapes cut to various dimensions. In addition to elongated triangles like Darwin’s, she made right-angled triangles and even four-sided figures. On the whole, she found that the sharpest angle was the one most often inserted in the worm’s burrow, and the shortest side the one least often drawn down by the worm.
| ||

Hanel then proposed a complex hypothesis to explain these results. An automatic reaction was involved, she held, stimulated by the relative length of the sides of the paper as perceived by the crawling worm. A succession of perceptions—shorter side, angle, longer side, angle—stimulated the worm to a pulling response. Like Darwin, however, Hanel stayed indoors at night, and did not watch the worms at work.
Now, the Principle of Occam’s Razor states, with obvious common sense, that the first hypothesis to test is the one that satisfactorily explains all the available facts in the simplest fashion. Darwin did exactly this in setting up his initial triangle experiment: he was testing the hypothesis that the worm’s work was a matter of pure chance. The flaw in his work was that Darwin did not actually watch the mechanics of the drawing-in operation as it proceeded in his own garden. Hanel, too, who dashed the earthworm’s brief reputation for intellect, did so by examining the results of their work, but not the work itself.
Had either done so, each would have observed what Hermann Jordan. the naturalist, was able to see with the aid of an oil lamp in 1913. For not in a single instance among the annelids that Jordan watched by night did any one of the worms crawl all the way out of its burrow! So much, then, for Hanel’s hypothesis of an elaborate stimulus resulting from the worm’s circumnavigation of the object to be seized. What about Darwin’s initial (and abandoned) hypothesis of trial-and-error attachment?
As Jordan watched, a worm’s front portion would suddenly appear from its burrow, rotate slowly in all directions and then grasp the first object with which it came in contact. This was the very
|
||

Now, most of the time the worm’s first tug would be unsuccessful: the leaf would catch itself in a position that straddled the burrow’s opening. At this point, Jordan found, the worm would either give up and withdraw or, letting go momentarily, would reattach itself to another spot on the leaf. Then, in these cases, through chance and perseverance, a point near the leaf’s apex might eventually be got hold of. The mechanics of the situation then permitting, success would attend the worm’s efforts.
Jordan’s nocturnal fieldwork dissipated both the aura of mystery that attached to Darwin’s statistics on purposeful leaf-pulling by the earthworm and, at the same time, destroyed Hanel’s hypothesis of complex reflex chains. We are left with a trial-and-error manner of procedure, the possibility of which Darwin had realized but which the great naturalist had felt justified in dismissing.
Lest, at this point, Lumbricus seem reduced to utter aimlessness, let us not forget that the earthworms’ "purposeless" actions could not be so labeled were it not for the repetitive "perseverance" which often impels a worm to let go and then attach itself again and again. The essence of this particular phenomenon—which may be dramatically proclaimed the "vital urge" or, more soberly, the "reactivity of the organism"—remains as much in the realm of conjecture as ever.
But the story does not end here. Jordan’s 1913 observations were submitted to a rigorous analysis by Otto Mangold, the German physiologist, in 1925. He amply substantiated Jordan’s conclusions, and added a number of additional—and intriguing—points.
Mangold was able to establish the correctness of one observation by Hanel—a certain amount of chemical discrimination exists in the earthworm, so far as preferred points of attachment to various leaves are concerned.
| |
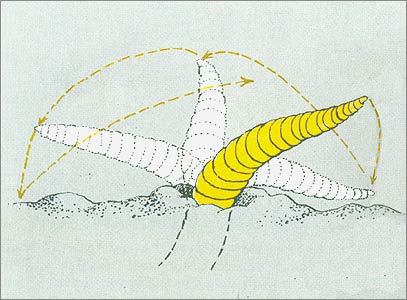

To demonstrate this, Mangold worked very ingeniously. First, he took wetted-down cherry tree leaves and, removing the stems, rolled them into narrow, cigarette-shaped cylinders. In these cylinders, one end consisted entirely of leaf tips and the other was composed of basal portions only. Out of fifty such plugs pulled into earthworm burrows, Mangold found that half had been drawn in by the “base” end and half by the “tip” end. These results seem to eliminate any possible chemical discrimination so far as different parts of the stemless leaf were concerned.
But what about stems? Mangold took pine needle bundles that had been completely deodorized by soaking in a strong acid-alcohol solution. These bundles were then dipped half their length into a gelatinous suspension composed of pulverized stems. After this coating had hardened, the other half of each bundle was similarly soaked with ground-up leaf suspension. Some twenty of these cylinders (with equally divided outer coatings of different composition) were set out: only one was drawn down by the “stem-half” thus demonstrating the earthworms’ clear-cut “preference” for the “leaf” chemicals over the “stem” ones.
| ||||
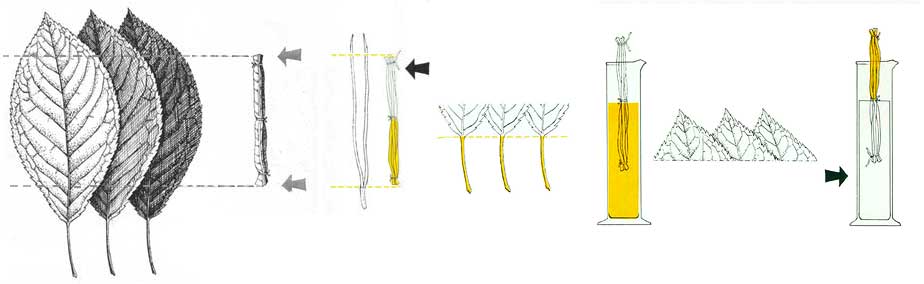
THE case for an initial random method of procedure—together with mechanical and chemical elements—now being more or less complete, there remains another aspect of earthworm behavior open to question. Is it possible for this invertebrate to profit from previous successful experience? Yerkes, perhaps best known for his work on anthropoid apes, studied this aspect of earthworm behavior early in his career (1912). Using a laboratory maze, he demonstrated that worms possess both some sort of memory and the ability to modify their behavior in terms of previous experience. Yerkes’ apparatus was a T-shaped passage: the floor of one arm was lined with sandpaper, beyond which were situated a pair of electrodes that would shock an advancing worm. The T’s other arm led to an artificial burrow, while the open stem was directed toward a light. To avoid the light, the worms moved toward the crosspiece of the T, where a choice presented itself: one “decision” led over very rough ground to a literally shocking experience; the other to a cozy burrow.
|
||
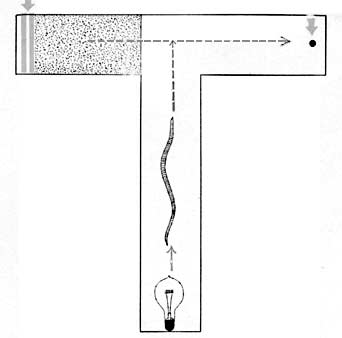
Yerkes’ worms, through repeated trials, avoided the sandpaper “decision” increasingly, and also avoided the shock—“warned” by the rough floor. In addition, early tendencies to retrace a path down the stem of the T, or to turn back after progressing well toward the “reward” burrow, disappeared. However, a hundred trial runs through the maze were required before the earthworms evidenced any benefit from their experience.
A later investigator, Rudolf Malek (1926), tested the annelids in Prague for associative learning ability in a more natural environment and found that, in their normal surroundings, earthworms seemed capable of learning much more quickly, than would have seemed possible on the basis of Yerkes’ results. Malek worked with worms that had partly emerged from their burrows and were rotating their upper bodies—“searching.” He offered them leaves that he held firmly in his fingers. Such fixed leaves were first gently seized by the annelid, then gripped harder three or four times in the same place in rapid succession. Then, exploring the leaf between attempts, the worms would try ten to twelve times at other points. After this, the animal abandoned the task, showing no further interest in that particular object.
It is not fatigue that causes the worm to desist. It will fasten onto another object at once after having abandoned the previous one. Indeed, a renewed attempt is made if the same leaf is offered in a place only an inch or so removed from the first. But a localized immovable object will not be tried again, although the memory thus indicated only lasts for a short time.
SOME evidence exists, then, for a limited learning ability in Lumbricus. However, the main operation we have been examining—the apical drawing-in of the leaf—is accomplished mostly as a chance effect. There is, thus, a certain irony in the history of earthworm behavior studies. Darwin, one of the first scientists to regard seemingly purposeful changes merely as the remnant of many random effects, screened out by natural selection, felt justified in dismissing this same possibility as applying to the activities of the lowly annelid.
Indeed, the crux of Darwin’s renowned theory of the origin of species, lies in its demonstration that the marvelous and precise "hand-in-glove" fit of organisms to their environment is not purposeful adaption to the situation but, rather, the end result of a long process of elimination—"nature" acting as the filtering agent.
Out of a constant stream of randomly occurring hereditary variations, only those which give the organisms possessing them some sort of biological advantage, compared to others
|
||
Thus, there is a selective premium on certain genetic differences: since they enable the possessor to live “better” and thus produce more young, they can be passed on more frequently to more descendants than hereditary contributions of lesser value in the environmental situation. Eventually, the greater portion of the existing population will carry more and more of those traits which make for being “well-adapted.”
In the ease of the earthworm, the situation is really no different, procedurally. Here, too, the “natural selection” imposed by the diameter of the burrow and the shape of the leaf acts to is weed out most of the varying and random modes of attachment—only those are successful which happen to “fit” the environment. That, in the majority of eases, the successful method happens to be pulling down by the leaf’s apex, certainly offers no surprise. Thus, the seeming puzzle that attracted Darwin’s attention is, at last, quite simply explained in terms of Darwin’s own grand scheme of variations rendered adaptive through the filtering process imposed by the environment.
Copyright © Natural History Magazine, Inc.