Featured Story
![]()
July-August 2006
|

The Scaly Ones
![]()
Squamata—lizards and snakes—have spread to almost every landmass and branched into more than 7,200 species. Ecological and molecular studies are bringing their family tree more clearly into focus.
![]()
By Laurie J. Vitt and Eric R. Pianka
They climb walls and scuttle upside down across ceilings, dive to the ocean floor to feed on algae, even glide through the air from treetops. Some, with no limbs and extremely long tails, look like snakes;
| ||
What particularly fascinates us is how, long ago, a common ancestor of squamates could have given rise to so many diverse descendants. Their living representatives—numbering more than 7,200 species—inhabit every continent except Antarctica and even many oceanic islands. When we look at them, we know that some of their similarities and differences reflect recent evolutionary adaptations, made in response to other species in their present-day environments. Other characteristics are a legacy of more ancient adaptations in response to unknown conditions—early choices that set various groups on separate evolutionary trajectories.
But with so many species and habitats, how can one reconstruct the group’s evolutionary history? Fossils tell only a limited story, and they are relatively rare. (Many of the ancestral species were small, and so their bones were less likely to be preserved—and more likely to be overlooked by paleontologists seeking bigger, more spectacular finds.) Investigators have barely begun to probe the genetic data that might bring us closer to understanding the evolutionary relations among living groups. But we think enough clues exist to sketch a coherent story.
Our story begins when squamates and their nearest relatives, the Rhynchocephalia (“beak-headed” reptiles, so named because their jaws have a beaklike tip) split from a common ancestor. According to Susan Evans, a paleontologist at University College London, the branching probably took place sometime during the
| ||

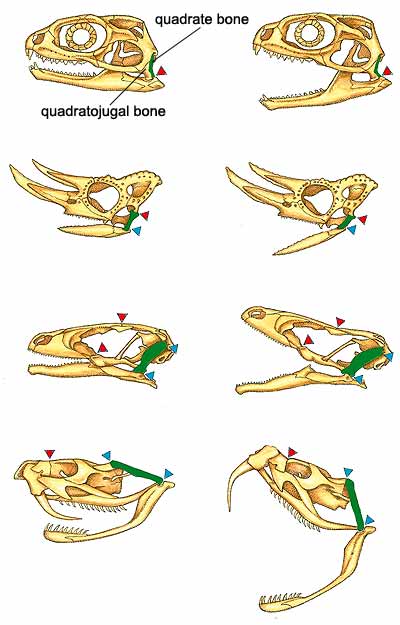
The common ancestor of squamates and rhynchocephalians was likely a small lizardlike reptile that ate insects and spiders. Although many skeletal features distinguish squamates from their sister group, one of the most important is found in the lower jaw. In the common ancestor, the lower jaw rotated from a pivot point on the bottom rear of the skull, which otherwise was relatively rigid. With the evolution of squamates, the skull bone that connected to the lower jaw—the quadrate—became only loosely attached by ligaments to the rest of the skull. That new hingelike configuration, known as streptostyly, enabled the back of the jaw to move more freely [see illustration below]. In practice, it enabled the animal to deliver a faster and more powerful bite, and perhaps to open its mouth wider as well, making it much easier for ancestral squamates to capture and handle prey.
Sometime between 30 million and 60 million years after the split between squamates and rhynchocephalians, the squamates themselves split into two major groups, Iguania and Scleroglossa. Fossil evidence for the timing of that split is sparse, but it is consistent with the amount of divergence evident in the DNA of living species—a question investigated by J. Robert Macey, a molecular biologist then at Washington University in St. Louis, and his colleagues. Intriguingly, the split appears to coincide with the breakup of Pangaea into two supercontinents, Laurasia and Gondwana.
|
For some time, paleontologists reasonably theorized that that event, by separating two populations of squamates, initiated the evolutionary divergence of the two major groups. Investigators now know that early fossils of both groups were deposited in both zones of Pangaea before the land mass broke up. Nevertheless, that geological event apparently bore some relation to the evolutionary one. Most of the early diversification of iguanians took place on Gondwana, the southern continent, whereas most of the early diversification of scleroglossans took place on Laurasia, the northern continent. Why that is, however, remains unclear.
When Iguania and Scleroglossa diverged, iguanians retained most of the characteristics of their squamate ancestors. Iguanian skulls remained relatively rigid. They continued the early squamate lifestyle of lying in wait for prey and remaining cryptic—hidden or camouflaged—except when pursuing prey. And they continued to rely on vision to detect and discriminate prey and on what Kurt Schwenk, a functional anatomist at the University of Connecticut in Storrs, calls lingual prehension—literally, holding with their tongues—to capture and manipulate prey. Schwenk argues that lingual prehension was the ancestral mode of feeding in tetrapod vertebrates, and so in squamates it may represent the inheritance of a very ancient trait.
|

The diversification of iguanians has been spectacular. Today they include most insect-eating lizards that tend to pursue their prey from fixed perches, such as chameleons; many other insectivorous species; and nearly all large plant-eating lizards around the globe—including iguanas, the group’s namesake. Among iguanians is Moloch, the thorny devil of Australia [see photograph left], a lizard that specializes in preying on ants and is so unusually shaped and well camouflaged that even experienced lizard biologists have difficulty spotting them.
Of the insect-eaters, chameleons have adopted the most extreme form of the ambush lifestyle. Chameleons have nearly eliminated pursuit; instead, they have a propulsive tongue that is often longer than their body [see photograph below]. A chameleon creeps within range of its insect prey, then hurls its tongue explosively at it. The prey sticks to the tongue, then gets rapidly pulled into the chameleon’s mouth. All the while, the chameleon remains otherwise immobile, hidden by its cryptic shape and coloration. One of the main advantages of that strategy is that it reduces risk: when an animal moves, potential predators may see it. The strategy also saves energy: shooting forth the tongue “costs” less than chasing down prey.
If there is a downside to the iguanian lifestyle, it is being tied down to real estate that matches the animal’s camouflage. A lizard that looks like a leaf can be nearly invisible in a bush, but it stands out on a rock. So the need for camouflage has doubtless played an important role in the evolution of iguanian social behavior. Most iguanians are territorial, fiercely defending areas where they blend in. There they remain unless another of their species usurps the favored place.
In sharp contrast to iguanians, scleroglossans evolved in ways that enabled them to capture and manipulate prey with their jaws, an ability that Schwenk calls jaw prehension. That strategy freed up the tongue for tasks other than capturing prey, and the tongue became firmer as a result (scleroglossa means “hard-tongued”). Among the early adaptations in this group was the addition of a joint in the top center of the skull that enabled the upper jaw to bend upward and downward (try doing that with your upper jaw!). The flexible movement in the middle of the skull, known as mesokinesis, greatly increased biting efficiency by allowing the upper jaw to move downward as the lower jaw closes from the bottom up [see illustration above]. Snakes have a similar hinge, but it is in front of their eyes (a condition known as prokinesis). They also have additional flexible joints, such as one at the front of the muzzle that enables the left and right bones of the upper jaw and palate to move independently (that is what makes it possible for a snake to “walk” its head down over a large item of prey). Another important innovation in snakes is a loose, stretchable cartilaginous ligament that joins the front tips of the right and left lower jawbones: that allows the bones to move apart, enabling snakes to swallow exceedingly large prey (recall the drawing of a snake in The Little Prince).
|

Switching from tongue to jaws for capturing prey might not seem groundbreaking. The change freed the tongue for other functions, however, and likely set the stage for the evolutionary enhancement of the vomeronasal organ, a chemosensory organ embedded in the roof of the mouth. That organ detects heavy, nonairborne molecules taken in through the mouth. It supplements nasal olfaction, which is the ability to smell airborne molecules that enter through the nostrils as the animal breathes. It is distinct as well from taste, which is the ability to analyze chemicals with taste buds on the tongue. In scleroglossans the enhancement of both olfaction and vomerolfaction was life-changing: not only could they rely less on sight to detect prey, but they could also perceive sex and sexual receptivity in their own species on the basis of chemical cues alone.
The avid way Gila monsters hunt out eggs suggests that enhanced chemical senses have helped some scleroglossans to find prey particularly rich in energy. But perhaps more important, the enhanced sense of vomerolfaction also enabled scleroglossans to identify dangerous prey—prey whose metabolic poisons or other defensive chemicals could be deadly or at least costly to digest. Several years ago we combined data we had gathered independently over our careers and demonstrated that, compared with iguanians, scleroglossans eat many fewer ants, other hymenopterans, and beetles, most of which produce noxious or toxic chemicals. Most scleroglossans are highly active, and so they encounter numerous potential prey; that behavior may afford them the luxury of passing over noxious insects.
As evolving chemical senses enabled scleroglossans to locate prey and potential mates without relying on vision, they adopted a more mobile searching strategy. This made them more conspicuous, requiring wariness, speed, and agility—not just camouflage—to escape predators. For the most part they ceased devoting time and energy to defending territories. Their chemical senses also enabled some scleroglossans to enter subterranean microhabitats and others to shift to nocturnal habits, both unavailable to iguanians that rely on vision as their primary sensory system.
One possible measure of the success of the shift to jaw prehension and chemical senses is that in terms of extant species, scleroglossans outnumber iguanians 4.5 to 1. That ratio might overstate an advantage resulting mostly from chance. For example, scleroglossans could have split into separate groups a bit earlier than iguanians did in their evolutionary history, and the present-day diversity might simply magnify that small advantage. But scleroglossan success was real. A more convincing measure is that scleroglossan species outnumber iguanian species at nearly every site where both occur. In most places, scleroglossans dominate the ground, foraging freely over the landscape, whereas most iguanians are confined to rocks, shrubs, tree trunks, or other more restricted habitats.
|
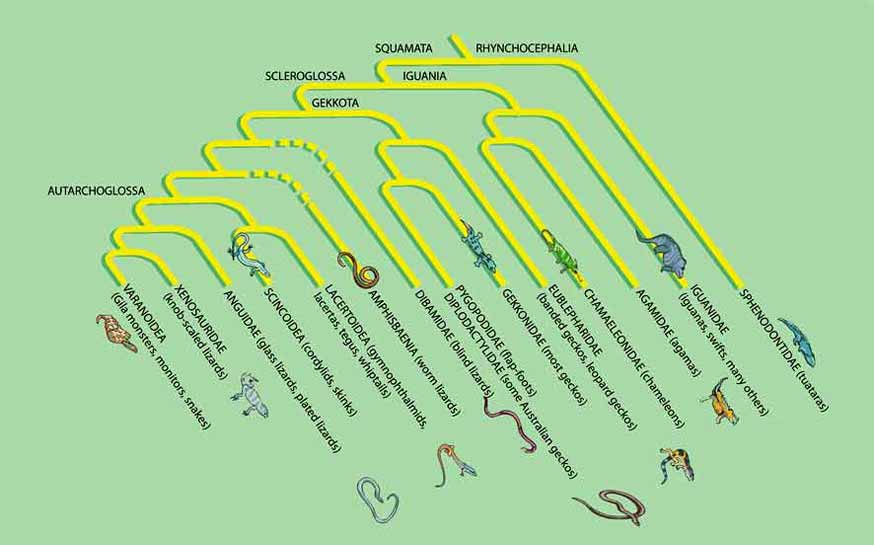
Subsequently—about 180 million years ago—scleroglossans split into two groups, Gekkota and Autarchoglossa, which differ in when they are active and how they use their tongues. Gekkotans include, among others, geckos (after which the group is named) and the snakelike flap-foots of Australia. Most geckos are nocturnal. They have no eyelids; instead, the eye is covered by a “spectacle,” or transparent scale, similar to the eye-covering of snakes. The tongue serves as a windshield wiper to clean the spectacles (as well as the lips). Geckos also have a good sense of smell. William E. Cooper Jr., a biologist at Indiana University–Purdue University in Fort Wayne, has shown that geckos detect airborne pheromones of other geckos, and discriminate prey on the basis of chemical signals.
|

Geckos are also noted for being able to scale vertical surfaces and walk upside down on leaves, rock faces, and trees. They can even jump from tree to tree by catching a leaf on the second tree with a toe. Although some other lizards (anoles and a few skinks) can climb vertical surfaces with their toe pads, none are as adept as the geckos. Kellar Autumn, a biologist at Lewis & Clark College in Portland, Oregon, has shown that gecko feet stick to surfaces by van der Waals forces, a form of molecular attraction. Their toe pads have ridges, each ridge covered with thousands of setae, hairlike structures that branch into hundreds of microscopic endings, called spatulae, that maximize contact with a surface.
Flap-foots belong to the Australian family Pygopodidae. Pygopodids are elongate lizards with no forelimbs and greatly reduced hind limbs. Some are burrowers, others terrestrial or arboreal. Many flap-foots swim through grass tussocks. Some are nocturnal, others diurnal. Some mimic venomous snakes in coloration or by acting menacingly.
One flap-foot, Lialis, acts more like a snake than it does other lizards in that it swallows very large prey, including other lizards. Skinks, Lialis’s primary prey, are armored with bony plates known as osteoderms embedded in their scales, and so, like some skink-eating snakes, Lialis has evolved hinged teeth. When a tooth hits an osteoderm, it folds, whereas a tooth that goes between scales remains erect, giving it good purchase. As the skink squirms and wiggles in the jaws of Lialis, it literally ratchets itself down the predator’s gullet.
Autarchoglossans, “independent tongue” reptiles, pick up heavy, nonairborne chemicals from surfaces with their tongues. The tongue then deposits the chemicals on the vomeronasal organ. Among the autarchoglossans are tegus and whiptail lizards, which march around in the hot sun in deserts and open tropical habitats of the New World,
|

The long, thin tongues of monitor lizards and snakes take chemical sensing to its most sophisticated level. Not only can they discriminate chemicals, but by comparing chemical dosages picked up on each fork of the tongue, they can detect edges as well as the direction of a chemical source. Just as chameleons are the logical endpoint to the evolution of a sit-and-wait foraging strategy, snakes might be considered the logical endpoint within a group of organisms that have highly flexible skulls and a well-developed chemosensory system for locating and discriminating prey.
And just as scleroglossans seem to have elbowed iguanians out of terrestrial habitats, so too autarchoglossans seem to have pushed gekkotans off the ground—or at least into a nocturnal lifestyle. Although one group of autarchoglossan lizards, the snakes, includes many nocturnal species (most from warm regions), autarchoglossans are generally active by day. In fact, their high activity levels usually require a high body temperature. Gekkotans operate at lower body temperatures, but their nocturnal habits have limited their distribution to regions that are warm enough at night. Most geckos, for instance, live in the tropics or in deserts, and only a few live in colder climates.
Autarchoglossans also continued another tradition of their scleroglossan ancestors. Whereas chemical senses had enabled the scleroglossans to expand their ranges and relax their need to defend territory where they were camouflaged, the continuing evolution of those senses enabled the autarchoglossans to forage in places where they could not see. And in several families of autarchoglossan lizards, species evolved that dug burrows and lived underground.
|

In many cases the subterranean lifestyle led to the evolutionary loss or reduction of limbs and to shortened tails. In some species, eyes were reduced to simple light-detecting organs—the animals had essentially shifted to a world dominated by chemical signals. Other lineages evolved elongate bodies and tails, even as limbs were lost or reduced, giving rise to what are known in the United States as glass lizards (genus Ophisaurus). (The common term “glass lizard,” however, can refer to species in several families of lizards that have evolved independently on different continents.) Neither limblessness nor subterranean activity has ever evolved in iguanian lizards.
That brings us to snakes, which are characteristically limbless. Although they figure large in people’s lore and imagination, snakes are simply one evolutionary group (Serpentes) of scleroglossan lizards. They are closely related to lizards that belong to the family Varanidae, which comprises the Komodo dragon and other monitor lizards. They may have descended from a varanidlike ancestor, and a common assumption is that they evolved from some burrowing terrestrial lizard. Another theory, however, is that the first snakes were aquatic, and [the wording that follows corrects an error in the published text —Ed.] were members of a group that shared a common ancestor with the mosasaurs, an extinct group of large marine reptiles that were also closely related to the varanids [see “Terrible Lizards of the Sea,” by Richard Ellis, September 2003]. But even if terrestrial snakes descended from an aquatic ancestor, herpetologists generally agree that today’s species of aquatic snakes have all descended from terrestrial snakes.
What a snake can find is limited only by the size of crevice or hole that it can stick its head into; what a snake can eat is limited in size only by how far it can disarticulate one of the most flexible skulls known in vertebrates. As a result, snakes have become top predators, coming back to haunt such close evolutionary relatives as the lizard autarchoglossans as well as iguanians and gekkotans.
|

Many venomous snakes have brightly colored rings (as do Gila monsters) that warn potential predators that they are dangerous. A host of nonvenomous snakes have evolved color patterns that mimic those of venomous species, thereby taking advantage of the protection a venomous reputation affords against potential predators. People are among the many large animals that instinctively give snakes a wide berth. The threat of a venomous snake, not to mention the dangers posed by large constrictors, probably ingrained that instinct in our early mammalian ancestors.
Whether we are attracted or repelled by snakes and other squamates, we owe the group respect for its evolutionary success. Along with turtles and crocodilians, they are the reptiles that we see around us today. Dinosaurs, ichthyosaurs, plesiosaurs, and pterosaurs were impressive in their time, but all (except for birds) were long gone before humans came along, as much as we might fantasize about “lost worlds.” Squamates, on the other hand, watched the dinosaurs come and go. Chances are they will be around to watch humans exit as well.
 |
Copyright © Natural History Magazine, Inc., 2006